

Продуктивность транспирации транспирационный коэффициент.
_Транспирация
Транспирация характерна не только для растений, но и для животных, ибо позволяет и тем, и другим регулировать температуру тела, спасая их от перегрева.
Завершающей частью водного обмена растений является транспирация, или испарение воды листьями, то есть верхний двигатель тока воды в растении. Это явление с физической стороны представляет собой процесс перехода воды в парообразное состояние и диффузию образовавшегося пара в окружающее пространство.

Схема транспирации
Транспирация выполняет в растении следующие основные функции:
- это верхний двигатель тока воды,
- это защита от перегрева,
- это нормализация функционирования коллоидных систем клеток листа.
Транспирация характеризуется следующими показателями: интенсивностью, продуктивностью и коэффициентом.
Интенсивность транспирации – это количество воды, испаряемой растением с единицы листовой поверхности в единицу времени.
Сопротивление пограничного слоя зависит от ветра, при отсутствии ветра оно максимально, чем больше ветер, тем оно меньше.
Устьичное диффузионное сопротивление зависит от степени открытия устьиц.
Кутикулярное диффузионное сопротивление зависит от толщины кутикулярного слоя, чем она больше, тем больше сопротивление.
Продуктивность транспирации – это количество созданного сухого вещества на 1 кг транспирированной воды. В среднем эта величина равна 3 г/1 кг воды.
Транспирационный коэффициент показывает сколько воды растение затрачивает на построение единицы сухого вещества, т.е. этот показатель является величиной, обратной продуктивности транспирации и в среднем равен 300, т.е. на производство 1 тонны урожая затрачивается 300 тонн воды.
Очень важным моментом в процессе транспирации является действие абиотических факторов окружающей среды: влажности атмосферного воздуха и температуры воздуха.
Чем менее влажен атмосферный воздух, т.е. чем меньше его водный потенциал, тем интенсивнее будет идти транспирация.
При 100% влажности воздуха его водный потенциал равен нулю. Уже при снижении влажности воздуха на 1-2% его водный потенциал становится отрицательной величиной, а при снижении влажности воздуха до 50% показатель водного потенциала выражается отрицательной величиной порядка 2-3 сотен бар в зависимости от температуры воздуха.
При этом в клетках листьев показатель водного потенциала, как правило, выше нуля, поэтому диффундирование воды из межклетников в атмосферу наблюдается почти всегда.
Чем выше температура воздуха, тем выше будет и температура листа, при этом температура внутри клеток листа может быть на 10 о С выше, чем в атмосфере. Происходит нагрев воды, находящейся в листе, что также способствует процессу испарения.
Регулировка транспирация происходит в растении по двум механизмам:
- устьичная регуляция,
- внеустьичная регуляция.
Наиболее существенной является устьичная регуляция, которая определяется как некоторыми физическими закономерностями, так и влиянием ряда факторов внешней среды и внутренней биохимией клеток листа.
С физической точки зрения основой испарения из устьица является физический механизм испарения с ограниченных поверхностей очень маленькой площади. При этом имеет значение величина снижения упругости водяного пара ( F- f) и расстояние (l), на протяжении которого поддерживается эта разница, которая определяет градиент дефицита насыщения.
Применительно к испарению с площади круга формула скорости испарения принимает вид
где k — значение всех прочих факторов, определяющих скорость испарения, а R — радиус круга.
При испарении с малых поверхностей, когда доля участия краевого испарения значительна, формула видоизменяется в
где n — положительное число между 1 и 2, т.е.2 > n> 1. В случае малых площадей, таких как отверстие устьичной щели, n становится равным 1. Таким образом определяющим становится фактор k, т.е. суммарное значение факторов окружающей среды и суммарное количество устьиц на листе.
В устьичной транспирации ведущими факторами являются:
- количество устьиц на единицу листовой поверхности,
- форма листа (чем более причудлива форма листа, тем больше его площадь, а, значит, и количество устьиц),
- наличие ионов К + (чем выше концентрация, тем больший приток воды в замыкающие клетки устьица и тем шире устьичная щель),
- наличие абсцизовой кислоты (чем выше концентрация этого гормона старения, тем меньше раскрытие устьица) (пример — мутант томата wilty),
- концентрация углекислого газа в подустьичной полости (чем ниже концентрация, т.е. меньше 0,03%, находящихся в воздухе, тем больший приток воды в замыкающие клетки устьица и тем шире устьичная щель),
- наличие солнечного света (на свету крахмал превращается в простые сахара, т.е. концентрация клеточного сока выше, поэтому наблюдается больший приток воды в замыкающие клетки устьица и раскрытие устьичной щели),
- наличие и скорость ветра (непосредственно к испаряющей поверхности прилегает слой воздуха, в котором водяной пар постепенно испаряется далее в атмосферу, при этом в безветренную погоду скорость испарения выражается линейной зависимостью между дефицитом насыщения воздуха и расстоянием от испаряющей поверхности. Однако, при наличии ветра, который » сдувает» испаряющиеся молекулы воды, происходит увеличение дефицита насыщения воздуха. Возле поверхности листа сохраняется лишь небольшой ламинарный слой (dS), сохраняющийся и при сильном ветре, где можно наблюдать линейную зависимость дефицита насыщения от расстояния).
Внеустьичная транспирация определяется количеством и размерами межклеточных пор в кутикуле листа. Радиус клеточных пор очень мал, составляет около 100-200 А о , т.е. около 0,00001мм, однако в листе имеющем много кутикулярных пор скорость испарения снижается достаточно значительно, иногда почти в два раза.
Различают три вида движения устьиц (закрытие и открытие устьиц):
- фотоактивные (под действием солнечного света),
- гидроактивные (при потере воды),
- гидропассивные (при дожде из-за набухания клеток эпидермиса и сдавливания устьичных клеток).
Суточный ход транспирации у всех растений определяется максимальной транспирацией в утренние часы и минимальной — в полуденные. При этом весьма существенное значение имеют и такие факторы, как температура почвы и воздуха, влажность почвы и воздуха, интенсивность солнечного излучения, наличие ветра.
Сезонный ход транспирации у многолетних растений определяется фазами развития растения.
Водный баланс в растении.
Водный баланс в растении поддерживается тогда, когда скорость поглощения воды равна скорости ее испарения. Обычно водный баланс в растении меняется в течение суток, при этом он зависит от уровня агротехники при выращивании растений, т.е. от уровня орошения и удобрения. Несбалансированность поступления и испарения воды проявляется в наличии водного дефицита, который наблюдается, как правило, у растений днем и отсутствует ночью.
В практике сельского хозяйства используются приемы, снижающие водный дефицит у растений: Использование освежительных поливов, Использование антитранспирантов.
Антитранспиранты делятся на две разновидности:
- вещества, вызывающие закрытие устьиц (абсцизовая кислота, фенилмеркурацетат),
- вещества, образующие пленки на листьях (полиэтилен, латекс).
Вот информация о Транспирационных коэффициентах некоторых культур:
Транспирационные коэффициенты, как показали наблюдения, в сотни раз превышают вес сухого вещества растения и колеблются в низких пределах.
Таблица 1. Транспирационные коэффициенты различных сельскохозяйственных культур
Транспирационный коэффициент и коэффициент водопотребления, зависимость от внутренних и внешних условий, способы их снижения;
Эффективность использования воды растением выражается рядом показателей. Количество созданного сухого вещества на 1 литр транспирированной воды характеризует продуктивность транспирации. В зависимости от условий выращивания и видовых особенностей растений она составляет 2-8, чаще 3-5 г/л. Величиной, обратной продуктивности транспирации, является транспирационный коэффициент, который показывает, сколько воды растение затрачивает на построение единицы массы сухого вещества. Транспирационные коэффициенты варьируют от 100 до 500.
Определить продуктивность транспирации или транспирационный коэффициент довольно сложно. Расчет потери воды на транспирацию за вегетационный период на основе данных об интенсивности транспирации по декадам или месяцам дает большую ошибку.
В полевых опытах и агрономической практике для оценки эффективности использования воды определяют коэффициент водопотребления (эватранспирационный), который рассчитывают как отношение эвапотранспирации к созданной биомассе. Под эватранспирацией понимают суммарный расход воды за вегетацию 1 га поверхности почвы (эвапорация) и транспирация.
Коэффициент водопотребления в значительной степени зависит от почвенно-климатических факторов. В засушливые годы он выше, чем в более влажные. Это объясняется тем, что в засушливых условиях усиление эвапотранспирации не сопровождается увеличением продуктивности растений, чаще она снижается, поэтому эффективность использования воды уменьшается.
Другим метеорологическим фактором, значительно влияющим на эффективность использования воды сельскохозяйственными растениями, является температура. С повышением температуры эвапотранспирация усиливается. Прохладный воздух снижает эвапотранспирацию, но у теплолюбивых культур вызывает также резкое подавление ассимиляционных процессов.
Мощным фактором снижения коэффициента водопотребления является повышение плодородия почвы. Снижение коэффициента водопотребления происходит не только при внесении удобрений, но и в случае любого изменения условий произрастания растений, сопровождающегося повышением урожая, в том числе и улучшения обеспечения их водой.
Водопотребление и урожайность связаны нелинейной зависимостью. При некотором достаточно высоком уровне урожайности ее рост уже не сопровождался повышением водопотребления, так как испарение в посеве или насаждении приближается к испарению со свободной водной поверхности.
Как оказалось, внешние условия не только регулируют степень открытости устьиц, но и оказывают влияние непосредственно на процесс транспирации. Зависимость интенсивности испарения от условий среды подчиняется уравнению Дальтона. Транспирация также подчиняется это формуле, правда, с отклонениями. Согласно уравнению Дальтона:
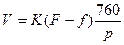
Где V- интенсивность испарения, количество воды, испарившейся с единицы поверхности; K- коэффициент диффузии; F- упругость паров воды, насыщающих данное пространство; f – упругость паров воды в окружающем пространстве при температуре испаряющейся поверхности; p- атмосферное давление в момент опыта.
Из приведенного уравнения видно, что испарение пропорционально разности (F-f), т.е. ненасыщенности атмосферы парами воды, или дефициту влажности. Чем больше дефицит влажности воздуха, тем ниже ее водный потенциал и тем быстрее будет испарение. Это в целом справедливо и для транспирации. Однако надо учесть, что при недостатке воды в листе вступает в силу устьичная и внеустьичная регулировка, благодаря чему влияние внешних условий сказывается в смягченном виде и транспирация начинает возрастать медленнее, чем это следовало бы, исходя из формулы дальтона. Несмотря на это, общая закономерность зависимости транспирации от насыщенности водой атмосферы остается справедливой. Чем меньше относительная влажность воздуха, тем выше интенсивность транспирации.
Сильное влияние на транспирацию оказывает свет. Если влияние влажности и температуры с большей силой сказывается на испарении со свободной водной поверхности, то свет сильнее влияет именно на транспирацию.
На интенсивность процесса транспирации оказывает влияние влажность почвы. С уменьшением влажности почвы транспирация уменьшается. Чем меньше воды в почве, тем меньше ее в растении. Уменьшение воды в растении автоматически снижает процесс транспирации в силу устьичной и внеустьичной регулировки.
Формула Дальтона выведена для спокойной погоды. Однако ветер, перемешивая слои воздуха, очень сильно увеличивает скорость испарения. Ветер оказывает влияние и на транспирацию, правда, по сравнению с испарением в несколько ослабленной форме. Поскольку ветер обычно не проникает внутрь листа, то под его влиянием возрастает в основном третий этап транспирации, т.е. перенос насыщенного водой воздуха от поверхности листа. В силу этого при ветре усиливается, прежде всего, кутикулярная транспирация. Большое действие ветер оказывает на транспирацию тех растений, где кутикула развита слабее. Сильнее на интенсивность транспирации сказываются суховеи. В этом случае ветер сгибает и разгибает листья и горячий воздух врывается в межклетники. Усиление транспирации уже на первом этапе.
Транспирация зависит от ряда внутренних факторов, прежде всего от содержания воды в листьях. Транспирация изменяется в зависимости от концентрации клеточного сока.
Транспирация изменяется в зависимости от величины листовой поверхности, а также при изменении соотношения корни/побеги. Чем больше развита листовая поверхность, больше побеги, тем значительнее общая потеря воды.
Интенсивность транспирации зависит и от фазы развития. С увеличением возраста растений транспирация падает.
Смена дня и ночи, изменение условий в течение суток наложили отпечаток и на процесс транспирации.
Что касается суточного хода транспирации, то в ночной период суток транспирация резко сокращается. Это связано с изменением внешних факторов , так и с внутренними особенностями. Измерения показывают, что ночная транспирация составляет всего 3-5% от дневной. При частом измерении транспирации можно заметить, что этому процессу свойственно ритмичное увеличение и уменьшение интенсивности. По-видимому, это связано главным образом с колебанием содержания воды в растении. Увеличение транспирации приводит к уменьшению содержания воды, это, в свою очередь, сокращает интенсивность транспирации. Как следствие, содержание воды растет, и транспирация также возрастает, и так непрерывно.
10. Методы учёта транспирации. Единицы измерения транспирации: интенсивность, экономичность, продуктивность транспирации, относительная транспирация. Транспирационный коэффициент.
Методы измерения интенсивности транспирации растений подразделяют на две группы. Первую составляют методы, основанные на учёте массы исследуемого объекта через заданные промежутки времени или непрерывно. Метод быстрого взвешивания отчлененного от растения листа или побега. Первое взвешивание проводят сразу после срезания, а второе — через 3—5 мин, что дает возможность измерять транспирацию при том состоянии насыщенности листа водой, в каком он находился на растении.
Методы второй группы основаны на учете величины потока водяного пара, поступающего из растения. В этом случае всю надземную часть растения или лист помещают в транспирационную камеру, через которую непрерывно прокачивают воздух. Пробы воздуха с входа и выхода камеры подаются в измерительное устройство, что позволяет регистрировать увеличение влажности воздуха в результате транспирации. Кол-во воды, испаряемой растением с единицы листовой поверхности в единицу времени, называют интенсивностью транспирации. Выражается в г/1м 2 или 1см 2 за 1час. Испаряемую воду можно отнести к массе листьев. Это также будет показателем интенсивности транспирации.
Продуктивность транспирации — количество граммов сухих веществ, образуемых при расходовании каждых 1000 г воды. Величиной, обратной продуктивности и транспирации, является транспирационный коэффициент, т. е. число граммов воды, израсходованной при накоплении 1 г сухих веществ. Интенсивность транспирации у большинства растений составляет 15-250 г*м2*ч днем и 1-20 г*м2*ч ночью. Продуктивность транспирации у растений в умеренном климате колеблется от 1 до 8 (в среднем 3 г) на 1000 г израсходованной воды, а транспирацинонный коэффициент — от 125 до 1000 (в среднем, около 300 т.е. около 300 г воды расходуется на накопление 1 г сухих веществ). Следовательно, на синтез веществ своего тела растение использует лишь 0.2% пропускаемой воды, остальные 99.8% тратятся на испарение.
Относительная транспирация – отношение воды, испаряемой листом, к воде, испаряемой со свободной водной поверхности той же площади за один и тот же промежуток времени. Экономность транспирации – количество испаряемой воды (в мг) на единицу (1кг) воды, содержащейся в растениях .Тонколистные растения расходуют за час больше воды по сравнению с растениями с мясистыми листьями, которые испаряют 8-20% от общего количества содержащейся в них воды. Транспирационный коэффициент показывает, сколько воды растения затрачивают на построение единицы сухого в-ва. Это величина, обратная продуктивности транспирации.
11. Особенности суточного хода движения устьиц у разных растении. Суточный ход процесса транспирации.
Английский исследователь Лофтфельд разделил все растения в отношении суточного хода устьичных движений на 3группы: 1.Растения, у которых ночью устьица всегда закрыты, утром устьица открываются, и их дальнейшее поведение в течении дня зависит от условий среды. Мало воды-они закрываются, достаточно воды-открыты. К этой группе относят прежде всего хлебные злаки. 2.Растения, у которых ночное поведение устьиц зависит от дневного. Если днем устьица были закрыты, то ночью они откроются, если днем были открыты, то ночью закроются. К этой группе относят растения с тонкими листьями-люцерна, грох, клевер, свекла, подсолнечник. 3.Растения с более толстыми листьями, у которых ночью устьица всегда открыты, а днем, как и у всех остальных групп растений, открыты или закрыты в зависимости от условий (картофель, капуста). Что касается суточного хода транспирации, то в ночной период суток транспирация резко сокращается. Это связано как с изменением внешних факторов (повышение влажности воздуха, снижение температуры, отсутствие света), так и с внутренними особенностями (закрытие устьиц). Измерения показывают, что ночная транспирация составляет всего 3-5%от дневной.
У деревьев, теневыносливых растений, многих злаков с совершенной регуляцией устьичной транспирации испарение воды достигает максимума до установления максимума дневной температуры. В полуденные часы транспирация падает и вновь может увеличиваться в предвечерние часы при снижении температуры воздуха. Такой ход транспирации приводит к незначительным суточным изменениям осмотического давления и содержания воды в листьях. У видов, способных переносить резкие изменения содержания воды в клетках в течение дня, наблюдается суточный ход транспирации с максимумом в полуденные часы. В обоих случаях ночью транспирация минимальна. Колебания интенсивности транспирации отражают изменения степени открытия устьиц в течение суток. Закрывание устьиц в полдень может быть связано как с увеличением уровня СО2 в листьях при повышении температуры воздуха (из-за усиления дыхания и фотодыхания, так и с возможным водным дефицитом, возникающим в тканях при высокой температуре, низкой влажности воздуха и особенно в ветреную погоду. Снижение температуры воздуха во второй половине дня способствует открыванию устьиц и усилению фотосинтеза.
12. Основные закономерности поступления воды в растение. Возникновение градиента водного потенциала в растении. Градиент водного потенциала как движущая сила водного тока в растении. Понятие водного потенциала, и его составляющие.
В почве вода поглощается корнями. Корень делится на четыре зоны: деления клеток, растяжения, всасывания, или корневых волосков, и проводящую, или опробковения. Зона деления клеток, защищенная корневым чехликом, нуждается в небольшом количестве воды. Водный потенциал определяется только матричными силами, т. е. способностью к набуханию коллоидов протоплазмы и клеточных стенок (у = ут). Интенсивное поглощение воды начинается с зоны растяжения. Здесь идет усиленное новообразование белков цитоплазмы. Поэтому водный потенциал определяется суммой матричного и осмотического потенциалов (у = уот + уп) и обеспечивает колоссальную способность поглощать воду. Зона корневых волосков является основной поглощающей зоной корня, которая направляет воду в русло дальнего транспорта. Здесь на 1 мм находится 230—500 корневых волосков (у = уп + ур). Проводящая зона корня характеризуется опробковением покровных тканей. Ее поглотительная функция заметно снижена, хотя показано, что опробковевшие части корня также могут поглощать воду.
Некоторое количество воды может поступать из атмосферы. Есть даже растения, для которых атмосферная влага является основным источником. К таким растениям относят прежде всего эпифиты, живущие на поверхности других растений, но не являющиеся паразитами. Они обладают воздушными корнями, в которых имеется многослойная ткань, состоящая из полых клеток с тонкими стенками. Такое строение позволяет им поглощать как парообразную влагу, так и воду осадков, подобно губке. У некоторых эпифитов дождевая вода собирается листьями и затем поглощается с помощью волосков. Приспособления к сбору дождевой воды листьями имеются и у других растений, например, у сем. зонтичные вода собирается в листовых влагалищах.
Водный потенциал выражает способность воды в данной системе совершить работу по сравнению с той работой, которую при тех же условиях совершила бы чистая вода. Водный потенциал определяют как величину, равную разности химических потенциалов воды в системе ( 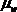 ) И чистой воды (
) И чистой воды (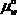 ) При той же температуре и том_же давлении, отнесенную к величине парциального молярного объема воды (
) При той же температуре и том_же давлении, отнесенную к величине парциального молярного объема воды (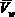 ), ψ =
), ψ =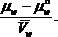 .
.
Водный потенциал, являясь фактически мерой активности воды, определяет термодинамически возможное направление ее транспорта. Молекулы воды всегда перемещаются от более высокого водного потенциала к более низкому, подобно тому как вода течет вниз, переходя на все более низкий энергетический уровень. Когда система находится в равновесии с чистой водой, у=0. В почве, растении, атмосфере активность воды и способность совершать работу ниже, чем у чистой воды, поэтому у обычно отрицателен. Водный потенциал имеет размерность энергии, деленной на объем, что позволяет выражать его в атмосферах, барах или паскалях.Водный потенциал растения является алгебраической суммой следующих четырех составляющих:
где уя — осмотический, |/;я — матричный, ур — гидростатический, у8— гравитационный потенциалы. Соотношение между ними и вклад в водный потенциал сильно различаются в зависимости от объекта и окружающих условий.
13. Верхний и нижний концевые двигатели водного тока. Гуттация и плач растений. Передвижение воды по растению. Апопласт и симпласт. Теория сцепления. Когезия и адгезия.
Концевые двигатели восходящего тока — нижний (корневое давление) и верхний (присасывающее действие транспирации) взаимосвязаны, а также имеют связь с другими процессами жизнедеятельности, что обеспечивается сложной системой эндогенной регуляции. Путь, который проходит вода в растении, можно разделить на две физиологически различные части: по живым клеткам и по проводящей системе. Радиальный транспорт воды в корне включает три системы: симпласт — совокупность протопластов всех клеток, соединенных плазмодесмами, апопласт, т. е. взаимосвязанную систему клеточных стенок и межклетников, а также дискретную систему вакуолей. Все они в большей или меньшей степени связаны с преодолением мембран. Даже путь воды по апопласту прерывается в эндодерме, где вода вынуждена проходить через живое содержимое пропускных клеток, из-за гидрофобных отложений в радиальных клеточных стенках, образующих пояски Каспари. Эта необходимость переключения на симпластический путь имеет большое значение, так как процесс передвижения по живой протоплазме может регулироваться в отличие от передвижения по клеточным стенкам. Следует отметить, что мертвые клетки обладают гораздо большей проницаемостью для воды, чем живые. Для нормального водообмена растения клетки корня должны оставаться живыми, их высокое сопротивление водному току вполне оправданно. Высокое сопротивление мембран, обладающих избирательной проницаемостью, позволяет корню контролировать ток воды в растении. Большая часть водного пути приходится на долю проводящей системы растения, состоящей из сосудов и трахеид. Установлено также, что вода и растворенные вещества могут передвигаться в стебле и в поперечном направлении. Это происходит, например, при подрезке корней или обрезке ветвей, в таком случае растение в состоянии обеспечить водой и веществами те части организма, которые раньше обслуживались утраченными органами. Возможность радиального транспорта обусловлена самой структурой проводящей системы, а именно: контактами сближающих трахеид через поры.
Примером работы нижнего концевого двигателя служит так называемый «плач» растений. Весной у деревьев с еще нераспустившимися листьями можно наблюдать интенсивный кселемный ток жидкости снизу вверх через надрезы ствола и даже верхних веток кроны. Явление «плача» свидетельствует о значительном корневом давлении, которое в этот период у основания ствола достигает не менее 10 атм. У вегитируюших растении при удалении стебля с листьями из оставшегося связанного с корнем пенька довольно долго выделяется ксилемный сок или пасока. Другим примером работы нижнего концевою двигателя является гуттация. При высокой влажности воздуха в результате деятельности нижнего концевого двигателя на концах и зубчиках листьев выделяется капельно-жидкая влага — растение гуттирует. Гутационное выделение влаги листьями особенно характерно для тропических растений, приспособленных к жизни в условиях повышенной влажности, при которых транспирация затруднена. В этих условиях подъем воды практически осуществляется только вследствие корневого давления. Под такими деревьями в тропическом лесу как будто постоянно идет дождь. Функцию выделения жидкости из тканей листьев выполняют специальные образования гидатоды. локализованные в зубчиках листьев.
Теория сцепления. Движущей силой восходящею тока воды в проводящих элементах ксилемы являемся градиент водного потенциала через растение от почвы до атмосферы. Он поддерживается двумя основными компонентами: 1) градиентом осмотического потенциала в клетках корня (от почвы до сосудов ксилемы), создаваемым активным транспортом ионов в живых клетках корня, включая молодые живые элементы ксилемы и 2) транспирацией. Поддержание первого градиента требует затрат метаболической энергии; на транспирацию используется энергия солнечной радиации. Градиент осмотическою потенциала обеспечивает поглощение воды корнем. Транспирация служит главной движущей силой восходящего тока воды.Согласно теории сцепления, вода в капиллярных трубках сосудов ксилемы поднимается вверх в ответ на присасывающее действие транспирации вследствие действия сил сцепления (когезии) молекул воды друг с другом и действия сил прилипания (адгезии) столба воды к гидрофильным стенкам сосудов. Обе силы препятствуют также образованию полостей у стенок сосудов, заполненных воздухом (или парами воды) и способных закупорить сосуд. При закупорке (эмболии) сосудов пузырьками воздуха всегда остается достаточное количество интактных нитей воды в других сосудах, чтобы обеспечить ток вверх. Кроме того. по-видимому, существуют механизмы восстановления непрерывности нарушенных эмболией водных нитей.
Источники:
http://ep-z.ru/posadki/ogorod/_transpiratsiya
http://studopedia.su/7_49983_transpiratsionniy-koeffitsient-i-koeffitsient-vodopotrebleniya-zavisimost-ot-vnutrennih-i-vneshnih-usloviy-sposobi-ih-snizheniya.html
http://studfile.net/preview/6726492/page:2/

